
| ЧЕЛОВЕК . ВОЗНИКНОВЕНИЕ РЕЧИ . СИСТЕМНЫЕ ИССЛЕДОВАНИЯ |
| ДОБРО ПОЖАЛОВАТЬ НА САЙТ ЮРИЯ ПАВЛОВА! |
|---|
|
|
|
|---|
|
|
Автоматизм и параллелизм обработки речевой информации мозгом. Информационная система
«Непрерывно пополняющийся комплекс данных указывает на
то, что самые ранние этапы фонологической, лексической, семантической и синтаксической обработки речи центральной нервной системой могут происходить автоматическим образом вне фокуса внимания. Анализ таких
независимых от внимания откликов на языковые стимулы показывает, что доступ к разнообразной лингвистической информации происходит в мозге достаточно быстро (в течение ~200 миллисекунд) и носит практически
одновременный, а возможно и параллельный характер».
Штыров Ю. Ю., Пулфермюллер Ф.
Автоматизм и параллелизм ранних этапов обработки речевой информации мозгом: нейрофизиологические данные на основе негативности рассогласования«Предисловие»
Обрабатывает ли наш мозг лингвистическую информацию различных типов одновременно или в определенном порядке? Требуется ли для этого наше внимание к речи или какие-то этапы ее обработки могут происходить автоматически? Как в психолингвистике, так и в психофизиологии нет однозначного ответа на эти вопросы. В настоящем обзоре мы рассмотрим ряд недавних исследований обработки речи мозгом человека, в которых использовалась т. н. негативность рассогласования (НР). Они показали, что, даже в условиях отсутствия внимания, характеристики этого мозгового отклика отражают физиологическую активацию следовых процессов памяти, связанных с различными элементами языка. Знакомые звуки родного языка вызывают более выраженную активацию, чем незнакомые; на более содержательном уровне, слова приводят к большей активации, чем бессмысленные словоподобные сочетания. Даже в отсутствие внимания к ним, такие словесные раздражители вызывают четкую последовательность активации коры головного мозга, которая начинается с верхних височных областей, а затем быстро распространяется в нижние фронтальные отделы левого полушария. Различные пространственно-временные паттерны такой корковой активности зависят от лексических и семантических свойств речевых стимулов, предоставляя таким образом разнообразные данные о конфигурации кодирующих такую информацию объединениях нервных клеток коры. На уровне более крупных высказываний возможна регистрация НР, отражающей соответствие предъявляемых сочетаний слов грамматическим правилам и семантическому контексту. Этот непрерывно пополняющийся комплекс данных указывает на то, что самые ранние этапы фонологической, лексической, семантической и синтаксической обработки речи центральной нервной системой могут происходить автоматическим образом вне фокуса внимания. Анализ таких независимых от внимания откликов на языковые стимулы показывает, что доступ к разнообразной лингвистической информации происходит в мозге достаточно быстро (в течение ~200 миллисекунд) и носит практически одновременный, а возможно и параллельный характер. ВведениеТрадиционные психолингвистические модели восприятия языка (Morton, 1969; Fromkin, 1973; Garrett, 1980; Dell, 1986; MacKay, 1987), находящие отражение и в разнообразных современных подходах к пониманию и производству речи (Levelt et al., 1999; Norris et al., 2000), различают несколько задействованных в этом процессе типов информации. Несмотря на обилие различий между этими теориями, они в целом соглашаются друг с другом в существовании: (1) уровня фонологической обработки, на котором (после примитивного акустического анализа) анализируются фонетические и фонологические характеристики звуков речи, (2) лексической обработки, иногда в виде концепции поиска информации в т.н. «ментальном лексиконе», т. е. своеобразном мысленном словаре, (3) семантической обработки, которая имеет отношение к восприятию конкретного смысла услышанного слова, (4) и наконец, синтаксического уровня, на котором идет анализ грамматической информации, обеспечивающей связь между словами в предложениях. В зависимости от конкретной теории, эти «модули» иногда объединятся, опускаются или увеличиваются в количестве. Однако, главный камень преткновения в современной психолингвистике заключается не в этом, а в разногласиях между моделями, предполагающими последовательный характер обработки этой информации, с одной стороны, и моделями, считающими, что все эти процессы происходят либо полностью параллельно, либо приблизительно одновременно, с другой. Этот вопрос может быть потенциально разрешен при помощи нейрофизиологии, которая, используя такие инструменты как электроэнцефалография (ЭЭГ) или магнитоэнцефалография (МЭГ), способна неинвазивно отслеживать мозговые процессы с большой временной точностью и регистрировать миллисекундные различия между ними. Нейрофизиологические эксперименты, действительно, сыграли заметную роль в этих дебатах. Разнообразные исследования, проведенные с использованием различного лингви- стического материала, сформировали уже устоявшийся взгляд на последовательный характер различных лингвистических процессов в мозге. После вычленения мозгом примитивных акустических характеристик из звукового сигнала (уже через 20- 50 мсек после начала стимуляции, напр., Krumbholz et al., 2003; Lutkenhoner et al., 2003), фонологический характер экспериментальных задач, как оказалось, способен изменять нервную деятельность только в диапазоне 100 - 200 мсек (Poeppel et al., 1996; Obleser et al., 2004). Обработка информации более «высокого» уровня, согласно некоторым исследованиям, требует от человеческого мозга значительно большего времени — компонент вызванных потенциалов под названием N400, имеющий свой максимум в диапазоне около 400 мсек после зрительного предъявле- ния словесного стимула (Kutas & Hillyard, 1980), обычно считается индексом семантических процессов. Иногда дополнительно выделяется его более ранний (350 мсек) субкомпонент, который, как предполагается, имеет отношение к лексическому доступу (Bentin et al., 1999; Embick et al., 2001; Pylkkanen et al., 2002; Stockall et al., 2004), что, кроме всего прочего, способствуют в целом спорному различению лексического и семантического уровней обработки. В случае синтаксических процессов, ситуация еще более усложняется. С латентностью около 100 мсек регистрируется т. н. ранняя левая фронтальная негативность (ELAN, early lef anterior negativity), которая увеличивается в ответ на некоторые грамматические нарушения в предъявляемых предложениях (Neville et al., 1991; Friederici et al., 1993). Дополнительно, в ответ на синтаксический материал регистрируются более поздние (>250 мсек) отклики, имеющие негативную полярность во фронтальных областях (Munte et al., 1998; Gunter et al., 2000). И наконец, одним из наиболее заметных синтаксических откликов является P600, поздний сдвиг вызванного потенциала, достигающий своей максимальной амплитуды (положительной полярности) около 600 мсек в центрально-теменных областях (см. обзор Osterhout et al., 1997; Osterhout & Hagoort, 1999). Такие данные, указывающие на последовательный доступ к различным типам информации, и сформировали доминирующую в нейрофизиологии языка и речи точку зрения (см. обзор Friederici, 2002). Она, однако, вступает в конфликт с целым рядом поведенческих данных, полученных в психолингвистических экспериментах. Речь идет об экспериментах, указавших на параллельный характер обработки мозгом информации о поступающих словах и о контексте их предъявления, и на быстроту этой обработки, которая может иметь место в течение первых ~200 мсек после того, как имеющееся информация позволяет распознать то или иное слово (Marslen-Wilson, 1973; Marslen-Wilson, 1987; Rastle et al., 2000; Mohr & Pulvermuller, 2002). Например, испытуемые могут совершать моторные ответы (нажатие кнопки) на зрительно предъявляемые слова в зависимости от своей оценки их фонетических и семантических свойств через 400 - 450 мсек после появления стимула (Marslen-Wilson & Tyler, 1975). Поскольку значительное время требуется на подготовку и выполнение самого акта моторного ответа, то наиболее ранние психолингвистические процессы, связанные с предъявленным словом, должны иметь место намного раньше. Еще более удивительным оказалось то, что даже в этом раннем диапазоне времени лингвистические процессы подвержены влиянию семантических и синтаксических факторов (Marslen-Wilson & Tyler, 1975). Более того, исследования с использованием методики повторения услышанного (shadowing) указали на то, что производство речи инициируется уже через 150?200 мсек после поступления языкового стимула (Marslen-Wilson, 1985). Ранние поведенческие эффекты, отражающие семантические процессы и контекстную интеграцию, были также получены в опытах с кросс-модальным праймингом, в которых было показано, что доступ к информации о предъявляемом слове может происходить задолго до его окончания, в течение 200 мсек после того, как имеющаяся информация позволяет его идентификацию, или даже раньше (Zwitserlood, 1989; Moss et al., 1997; Tyler et al., 2002). Эксперименты, использующие методику отслеживания движений глаз (eye-tracking) также показали, что ряд психолингвистических свойств предъявляемых стимулов оказывает влияние на движения глаз с такой же ранней латентностью (Sereno & Rayner, 2003). Исследования такого рода предлагают альтернативную точку зрения, согласно которой одновременный доступ к различным типам лингвистической информации может инициироваться в мозге в течение 200 мсек после поступления соответствующих раздражителей. Это, в свою очередь, ставит под сомнение последовательные модели обработки языка мозгом, предложенные в психолингвистике и поддержанные, по крайней мере частично, ранними нейрофизиологическими экспериментами. Другим спорным вопросом речевой науки является вопрос о том, насколько человеческая способность к обработке языка требует ресурсов других когнитивных систем, прежде всего внимания, и, наоборот, в какой степени она обладает автоматизмом. Та легкость, с которой мы можем воспринимать весь сложный комплекс поступающей в нашу слуховую систему речи, и простота, с которой мы, казалось бы, мгновенно расшифровываем заключающуюся в ней грамматическую, морфологическую, семантическую и прочую информацию, зачастую занимаясь при этом другими делами, привели к предположениям, что лингвистическая деятельность осуществляется мозгом в значительной степени автоматизированным образом (Fodor, 1983; Garrett, 1984; Garrod & Pickering, 2004; Pickering & Garrod, 2004). Чтобы рассмотреть этот вопрос в контексте настоящего обзора, мы разберем нейрофизиологические данные, полученные вне фокуса внимания с использованием пассивной экспериментальной парадигмы и мозгового отклика известного под названием «mismatch negativity (MMN)», чей русский перевод «негативность рассогласования (НР)» мы будем использовать в дальнейшем. НР — это мозговой отклик (отклонения вызванного потенциала или произведенного им магнитного поля) вызываемый редкими (т.н. девиантными) акустическими посылками, предъявляемыми изредка на фоне частых (т.н. стандартных) стимулов (Alho, 1995; Nааtаnen, 1995). Одной из важных особенностей НР является возможность ее генерации в отсутствие внимания испытуемых к слуховым раздражителям (Tiitinen et al., 1994; Schruger, 1996). В связи с этим считается, что она отражает автоматическую детекцию мозгом изменений в поступающей звуковой информации, являясь своего рода уникальным индикатором того, как наш мозг обрабатывает акустические события (Nааtаnen, 1995). В последнее время НР все больше и больше используется и для исследований нервных механизмов лежащих в основании речевой функции (Nааtаnen, 2001; Pulvermuller & Shtyrov, 2006). Предлагается четыре основных определяющих фактора: (a) НР является ранним мозговым откликом; (b) НР обладает автоматизмом; (c) она является откликом на индивидуальный раздражитель; (d) НР является откликом на перемену в стимуляции. Первые два момента относятся к известным свойствам НР, связанным тем или иным образом с механизмами ее генерации в мозге, а последние два пункта представляют из себя важность в связи с техническими, методологическими особенностями исследований активности мозга в ответ на языковые стимулы. Ранняя латентность НРКак уже говорилось, ряд поведенческих исследований указывает на запуск психолингвистических процессов в мозге в течение 200 мсек после поступления достаточной для этого речевой информации. Если эти процессы находят отражение в динамике событийно связанной мозговой активности, то соответствующие мозговые отклики, регистрируемые на поверхности головы, должны иметь место в том же диапазоне времени или даже раньше. Это является обязательным (хотя и недостаточным) условием для установления их прямой связи с вышеуказанными лингвистическими процессами; в противном случае, они могут быть всего лишь отдаленными последствиями этих процессов. Это автоматически исключает поздние сдвиги вызванных потенциалов (ВП; N400, M350, P600) из рассмотрения в качестве потенциальных кандидатов в индикаторы ранних языковых явлений в мозге.
Ранние, т. н. облигаторные слуховые потенциалы (P1, N1) не обнаружили чувствительности к лингвистическим факторам. ELAN, ранний синтаксический отклик, вполне соответствует временным требованиям, но не регистрируется в большинстве лингвистических задач (и более того, может быть найден далеко не во всех синтаксических опытах). С другой стороны, НР, во-первых, является ранним компонентом (обычная латентность в пределах 100 - 200 мсек), а во-вторых, прочно связана с такими когнитивными явлениями как память, внимание и его ориентирование, примитивный интеллект слуховой системы (Nааtаnen, 1995; Nааtаnen & Alho, 1995; Nааtаnen, 2000; Nааtаnen et al., 2001). Известно, что НР обладает чувствительностью к крайне абстрактным свойствам звуковых сигналов и таким образом является потенциальным кандидатом для идентификации предполагаемой ранней языковой деятельности в мозге.
Автоматизм НРХотя вопрос о том, подвержена ли НР влиянию процессов внимания, и если да, то до какой степени, нерешен, неопровержимо то, что НР легко вызывается в отсутствие прямого внимания к стимуляции. Таким образом, НР не требует активного участия испытуемого в осмыслении предъявляемого материала; как правило, испытуемый занят не связанной со слуховыми стимулами задачей (напр., просмотр фильма), в то время как НР, тем не менее, генерируется редкими случайными изменениями в поступающих слуховых сигналах. По крайней мере в этом смысле НР может считаться автоматическим мозговым откликом. Это, в свою очередь, имеет важное значения для ее использования в речевых исследованиях. Обычно в языковых экспериментах испытуемым дается задание внимательно воспринимать предъявляемые им слова или предложения (напр., Neville et al., 1991; Osterhout & Swinney, 1993; Friederici et al., 2000). Зачастую, инструкция включает в себя проведение какой-либо оценки лингвистического материала (напр., знакомые-незнакомые, правильные-неверные слова) вплоть до сложных лингвистических задач — таких как оценка грамматики предложения или лексическое решение (lexical decision). Поскольку в таком эксперименте требуется активное внимание со стороны испытуемого, невозможно удостовериться в том, что регистрируемая нейронная активность не связана скорее с мозговыми коррелятами процессов внимания, чем с языковыми процессами как таковыми. Связанные с процессами внимания явления, как известно, модулируют ряд вызванных откликов, в том числе связанных с мозговыми структурами находящимися вблизи основных языковых областей или даже перекрывающимися с ними (см. напр. Picton & Hillyard, 1974; Alho, 1992; Woods et al., 1993a; Woods et al., 1993b; Tiitinen et al., 1997; Escera et al., 1998; Yamasaki et al., 2002; Yantis et al., 2002). Вполне возможно, что испытуемые уделяют больше внимания необычным или содержащим преднамеренные ошибки стимулам (в подобных экспериментах зачастую используются псевдослова, бессмысленные предложения и грамматические ошибки) в то время, как они пытаются осмыслить их; возможно, что они применяют различную стратегию при обработке правильно сформированных и намеренно ошибочных стимулов. Такие специфические изменения стратегии и вариации в уровне внимания могут, в свою очередь, найти отражение в измеряемой активности, перекрываясь с подлинно языковой деятельностью мозга. Таким образом, крайне важным представляется ограничение влияния процессов внимания и эффектов специфики поставленной задачи. Для решения этой проблемы НР представляет собой идеальный инструмент, т. к. она регистрируется, когда испытуемые не концентрируют внимание на стимуляции и не выполняют никаких связанных со стимулами заданий.
НР как отклик на индивидуальный раздражительНесмотря на то, что наличие ранних психолингвистических процессов следует из имеющихся поведенческих данных, нейрофизиологам в основном не удалось найти мозговых коррелятов этой деятельности. Одна из возможных причин этой неудачи может иметь методологическую природу. В большинстве экспериментов по физиологии речи исследуются и сравниваются друг с другом большие группы стимулов; усредненные отклики при этом используются для обобщения результатов в применении ко всему материалу, относящемуся к той или иной категории лингвистических стимулов. Это приводит к проблеме физической вариативности стимулов, так как раздражители различаются по своим физическим свойствам (напр., длительность, спектральные характеристики, распределение звуковой энергии и т. п.). Как известно, даже минимальные различия в простых физических свойствах могут приводить к различиям в регистрируемой активности мозга (Nааtаnen & Picton, 1987; Korth & Nguyen, 1997), которые могут перекрываться во времени с лингвистическими эффектами, маскировать или скрывать их, или даже ошибочно казаться таковыми. Другой проблемой является психолингвистическая вариативность, т. к. стимулы, как правило, различаются по своим лингвистическим признакам, таким как частота встречаемости в языке или временные параметры распознавания слова. Последние при этом особенно трудно поддаются контролю, т. к. различные слова, даже при одинаковой общей длительности, поддаются распознанию и выделению из круга своих лексических конкурентов в различные моменты времени — в экстремальных случаях вскоре после начала слова или только чрез некоторое время после его окончания (Marslen-Wilson,1987). Несомненную пользу при этом может приносить традиционное выравнивание средних параметров между различными категориями стимулов в эксперименте, которое, тем не менее, по прежнему не решает всех проблем. Как правило, все известные ранние компоненты событийно-вызванных откликов имеют относительно малую амплитуду и короткое время проявления; если гипотетические ранние корреляты языковой деятельности обладают теми же свойствами, то вариативность внутри группы стимулов приведет к их размыву или полному исчезновению в среднем отклике по группе (Pulvermuller, 1999; Pulvermuller & Shtyrov, 2006). С другой стороны, более поздние отклики (такие как N400, P600), менее подвержены такому деструктивному воздействию процедуры усреднения, т. к. они обладают значительной амплитудой и длятся на протяжении сотен миллисекунд. Поэтому, чтобы обнаружить предполагаемые ранние эффекты, необходимо максимально уменьшить влияние вариативности раздражителей. Поскольку НР является откликом на один и тот же девиантный стимул, который для улучшения соотношения сигнала к шуму многократно повторяется случайным образом, то она предоставляет полное решение проблемы вариативности, сводя последнюю к нулю.
НР как отклик на изменениеПо своему определению, НР регистрируется как разница между откликами на девиантный и стандартный стимулы, контраст между которыми и приводит к ее появлению. Это, в свою очередь, дает уникальную возможность контроля над акустическими параметрами стимуляции в языковых экспериментах. Предлагаемый контроль состоит в использовании идентичных акустических контрастов в различных категориях стимулов, при одновременной манипуляции их лингвистическими свойствами. Рассмотрим следующие гипотетические сочетания раздражителей, используя английский язык, который применялся во многих наших экспериментах: (1) слово «ray» (луч, ед.ч.) в качестве стандартного стимула и «rays» (лучи, мн.ч.) — в качестве девиантного, (2) «lay» (класть, неопр.ф.)- «lays» (кладет, ед.ч. 3 л.), (3) «gray» (серый) — «graze» (пастись), (4) «tay» — «taze». Во всех четырех сочетаниях, НР будет вызываться одним и тем же акустическим контрастом, который состоит в наличии звука [z] в конце девиантного стимула в том положении, где стандартный раздражитель уже завершился. Таким образом, чисто акустическая НР в ответ на этот контраст должна остаться неизменной во всех четырех условиях. Однако эти четыре комбинации, несмотря на идентичность акустических контрастов, резко различаются по своему лингвистическому контексту: в первом из гипотетических экспериментальных блоков этот контраст представляет собой склонение существительного (изменение в числе); во втором блоке мы имеем спряжение глагола (в 3?е лицо). В третьем условии тот же самый акустический контраст приводит к полной лексической и семантической перемене — стандартные и девиантные стимулы не связаны общим смыслом и даже относятся к разным частям речи. Наконец, четвертая пара представляет из себя бессмысленные псевдослова, давая, таким образом, дополнительную возможность контроля над акустическими параметрами. Таким образом, записывая НР в ответ на такие акустически идентичные контрасты, можно сосредоточиться на эффектах различных лингвистических контекстов, не имея обычных ограничений, связанных с различием акустических параметров стимулов. Более того, возможно предъявление тех же самых девиантных стимулов в качестве частых стандартных раздражителей в отдельном экспериментальном блоке; в таком случае, НР может быть рассчитана как разница в отклике на физически один и тот же стимул в качестве девиантного и стандартного, что дает возможность еще более строгого контроля над физическими параметрами стимуляции. Использование строго контролируемых таким образом параметров стимуляции оказалось крайне успешным в ряде экспериментов. К их рассмотрению мы и перейдем в следующем разделе.
Экспериментальные данныеНа момент написания этого обзора (2008 год), имеющиеся экспериментальные данные показали чувствительность НР к языковым процессам, относящимся ко всем основным типам лингвистической информации: фонологическому, лексическому, семантическому и синтаксическому (рис. 1, 2).
Отдельные фонемы и их простые комбинации — слоги — были первым лингвистическим материалом, испытанным в парадигме рассогласования. Как показали эти эксперименты, звуки родного языка, например, гласные, вызывают более выраженную НР, чем их аналоги, не представленные в родной фонологических системе испытуемых (DehaeneLambertz, 1997; Naatanen et al., 1997). Важно, что это наблюдается вне зависимости от величины акустических различий между стандартными и девиантными стимулами. Более того, тогда как обычно НР имеет симметричную топографию, иногда с тенденцией к правополушарному преимуществу (Paavilainen et al., 1991), фонетическая НР имеет преимущественно левостороннее распределение, что предполагает возможную связь ее происхождения с языковыми областями коры, расположенных в левом полушарии головного мозга (Nааtаnen et al., 1997; Shtyrov et al., 1998; Shtyrov et al., 2000). Дальнейшие эксперименты показали, что изменения в паттерне НР могут отражать развитие новых фонологических представительств при освоении языковых навыков детьми или изучении нового языка взрослыми испытуемыми (Cheour et al., 1998; Winkler et al., 1999). Последующие опыты продемонстрировали чувствительность НР к фонотактическим факторам (Dehaene-Lambertz et al., 2000; Bonte et al., 2005; Bonte et al., 2007), паттернам ударения (Honbolygo et al., 2004; Weber et al., 2004), аудио-визуальной интеграции фонетической информации (Colin et al., 2002; Colin et al., 2004) и ряду других фонологических переменных. Время проявления этих фонетических и фонологических эффектов в НР варьируется от примерно 100 мсек (Rinne et al., 1999) до ~200 мсек (Shtyrov et al., 1998, 2000). Для объяснения этих результатов было предложено, что, в дополнение к детекции слуховых изменений и процессам кратковременной памяти, НР обладает чувствительностью к долговременным следам памяти, которые сформировались в нервной системе испытуемых в процессе их предыдущего взаимодействия с устной речью (Nааtаnen et al., 1997; Shtyrov et al., 2000; Nааtаnen, 2001). Из этого следуют важный вывод о том, что такие долговременные следы памяти могут активироваться предъявлением соответствующего девиантного стимула в парадигме рассогласования, и что эту активацию можно зарегистрировать нейрофизиологически даже в отсутствие внимания к стимуляции и какой бы то ни было связанной со стимулами задачи. Это послужило мотивацией к последующим экспериментам с использованием НР, которые ставили своей целью исследование лингвистических процессов более «высокого» уровня.
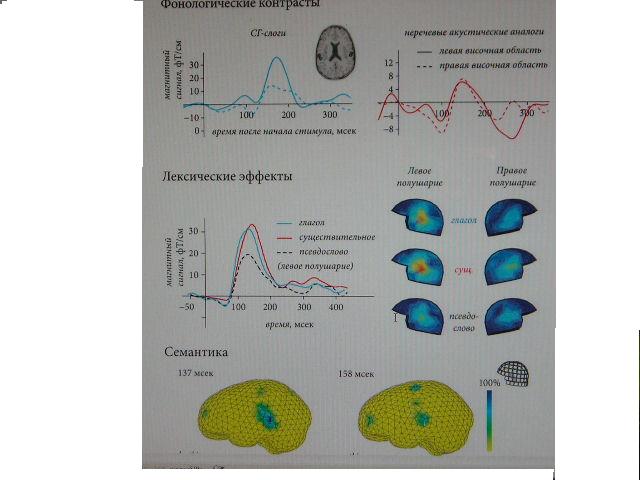 Рис. 1. НР как индекс обработки лингвистической информации мозгом: усиление НР в ответ на слоги родного языка (адаптировано из Shtyrov et al., 2000), лексическое усиление НР в ответ на значимые слова (адаптировано из Shtyrov et al., 2005), проявление семантических свойств слов в конфигурации источника НР (адаптировано из Pulvermuller et al., 2005).100 200 300 4 8 12 100 200 300 0 10 20 30 100 200 300 400 10 20 30 hotki potki 0% 100% Штыров Ю. Ю., Пулфермюллер Ф.
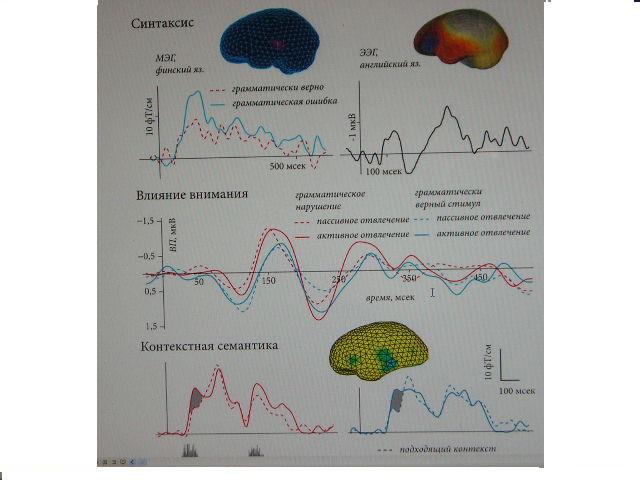 Рис. 2. НР в ответ на предъявление лингвистической информации «высокого порядка»: синтаксическая НР при использовании грамматически верных и ошибочных фраз (адаптировано из Pulvermuller & Shtyrov, 2003, и Shtyrov et al., 2003), ранний автоматизм и последующая подверженность процессам внимания в синтаксической НР (адаптировано из Pulvermuller & Shtyrov, 2008),НР и контекстно-семантическая интеграция (адаптировано из Shtyrov & Pulvermuller, 2007).1,5 0,5 450 50 150 350 250
Лексические процессыНаподобие того, как НР демонстрирует фонологическое увеличение в ответ на предъявление фонем и слогов родного языка, негативность рассогласования, вызванная отдельными словами, обладает большей амплитудой, нежели НР в ответ на подобные бессмысленные словоподобные (т.е. подчиняющиеся фонологическим принципам языка) стимулы (рис. 1). В проведенной нами серии исследований на эту тему мы предъявляли испытуемым наборы акустически сбалансированных слов и псевдослов и обнаружили увеличение амплитуды НР в тех случаях, когда в девиантной позиции находится значимое слово (Pulvermuller et al., 2001; Shtyrov & Pulvermuller, 2002; Pulvermuller et al., 2004; Shtyrov et al., 2005). Это усиление амплитуды, обычно имеющее свой пик в диапазоне 100 - 200 мсек, лучше всего объясняется активацией следов памяти для данных слов, реализованных в коре головного мозга в виде связанных между собой распределенных популяций нейронов (Pulvermuller & Shtyrov, 2006). Лексическое усиление НР было неоднократно подтверждено нашей группой и другими исследователями, причем в этих опытах использовались различные параметры стимуляции, методики регистрации откликов и языки (Korpilahti et al., 2001; Kujala et al., 2002; Sittiprapaporn et al., 2003; Endrass et al., 2004; Pettigrew et al., 2004). Как оказалось, для появления этого эффекта критическое значение имеет лексический статус именно девиантного стимула. В то же время мы не нашли эффектов лексического статуса стандартных раздражителей на амплитуду НР (Shtyrov & Pulvermuller, 2002), хотя в некоторых работах предлагается, что вызванные отклики на стандартные речевые посылки также могут реагировать на их лексический статус (Diesch et al., 1998; Jacobsen et al., 2004).
Исследования вызванной словесными стимулами активации рассогласования с помощью современных методик картирования мозга, таких как многоканальная магнитоэнцефалография (МЭГ) в сочетании с математическими алгоритмами оценки источников активности, позволили, в свою очередь, изучить связанные с восприятием слов мозговые процессы с недоступной прежде пространственно-временной точностью. Используя НР полученную в ответ на отдельные слова, мы смогли продемонстрировать, что эта активация сначала проявляется в верхних височных областях коры. Это происходит вскоре (~130 мсек) после того, как доступная информация позволяет идентификацию стимула. Затем, в строго определенном порядке (с задержкой около 20 мсек), она распространяется в нижние фронтальные области, что, возможно, отражает поток передачи информации в перисильвиевых языковых областях в процессе лексического доступа (Pulvermuller et al., 2003). Более того, мы обнаружили корреляцию между латентностью НР у индивидуальных испытуемых и индивидуальными моментами распознавания слова (word recognition points, WRP, см. MarslenWilson, 1987), определенными поведенческим путем, что непосредственно указывает на способность НР к отражению процессов распознавания слов и на связь между этими двумя явлениями (Pulvermuller et al., 2006). В результате непрерывного накопления данных такого рода, паттерн откликов рассогла- сования в ответ на девиантные словесные раздражители стал считаться своего рода индикатором, показывающим индивидуальные характеристики следовых процессов памяти для этих слов. Это, в свою очередь, дало возможность исследовать связанные со словами мозговые процессы без влияния со стороны механизмов внимания и вне зависимости от связанных со стимуляцией задач. Действительно, последующие исследования языковой функции с помощью НР помогли составить более полную пространственно-временную картину активации мозга, потенциально связанной с заключенной в словах семантической информацией.
Семантические процессыПервые эксперименты, использовавшие НР для изучения семантической обработки языка мозгом, были связаны с моделью, предсказывающей соматотопию (телорасположение) связанных с движениями слов (somatotopy of action words, Pulverm?ller, 1999; 2005). Эта модель является частью общего теоретического направления, в котором предполагается, что слова представлены в нервной системе в виде специфических следов памяти, существующих как распределенные сети нейронов. Последние формируются в результате усиления двусторонних связей между различными (в т.ч. удаленными друг от друга) областями мозга по мере того, как индивидуум получает опыт взаимодействия с движениями, объектами и концепциями одновременно с описывающими их словами (Hebb, 1949; Pulvermuller, 1999). Такая связь слова с его конкретным значением является неотъемлемой частью его семантики (Frege, 1980). Связанные с движениями слова, например, такие как описывающие движения нижних конечностей «бежать» или «пинать», или относящиеся к моторике рук «брать» или «рисовать», используются для описания движений тела. Таким образом, данные моторные действия являются сущностной характеристикой описывающих их слов (хотя вся семантика последних, возможно, этим и не исчерпывается). Если лексическое представительство такого слова проявляется в коре головного мозга в виде нейронной сети, то именно связи между группами нейронов в задействованных корковых областях (в данном случае языковых и двигательных) и должны реализовывать семантические взаимоотношения между внешней формой слова и описываемым им действием. Как известно, моторная кора организована соматотопически (Penfeld & Rasmussen, 1950), что в свою очередь ведет к выводу, что относящиеся к разным частям тела слова должны активировать, наряду с классическими языковыми областями коры, соответствующие части моторного неокортекса. Этот вывод, получивший подтверждение в ряде экспериментов с картированием мозга при помощи фМРТ и ЭЭГ (Hauk et al., 2004; Hauk & Pulvermuller, 2004), был также подвергнут проверке с помощью НР. При этом использовались различные методики записи НР (ЭЭГ, МЭГ) в совокупности с разными языками стимуляции (английский, финский). В этих экспериментах (Shtyrov et al., 2004; Pulvermuller et al., 2005), кроме обычно наблюдаемых источников активности в верхних височных областях, слова, относившиеся к движениям лица и рук, приводили к активации нижних и центральных областей фронтального неокортекса, включая, очевидно, корковое представительство верхней части тела. Более того, слова связанные с движениями нижних конечностей приводили к дорзальноцентральной активации, возможно связанной с сенсомоторным представительством ног (рис. 1). Эта специфическая дорзальная активация в ответ на связанные с движениями ног словами проявляется позднее (~170 мсек), чем более латерально распределенная активация в ответ на слова, связанные с движениями рук и лица. Такие пространственно-временные характеристики указывают на то, что источники НР в перисильвиевых областях, вкупе с почти одновременной активацией вдали от сильвиевой борозды, могут отражать доступ к смыслу слов в коре головного мозга человека. Минимальные задержки между локусами активности, возможно, обусловлены расстоянием и скоростью передачи потенциалов действия между задействованными областями коры. Эти данные указали на то, что обработка семантической информации, содержащейся в связанных с движениями словах, находит свое отражение в НР уже через 140 - 180 мсек после того, как акустический сигнал позволяет идентификацию слов. Следовательно, как и в рассмотренном выше случае лексического доступа и отбора, доступ к смыслу сказанного очевидно является ранним мозговым процессом, который также имеет место в рамках первых 200 мсек; таким образом, можно сделать вывод о том, что оба эти процесса происходят примерно одновременно. Не менее интересны результаты, полученные в опытах с НР и контекстной семантической интеграцией. В этих экспериментах целые предложения или комбинации слов предъявлялись испытуемым в пассивной парадигме рассогласования; в то же время их физико-акустические свойства находились под жестким контролем. Некоторые девиантные комбинации включали в себя семантический конфликт между словами, наподобие традиционной парадигмы семантических нарушений, обычно производящей отклик N400. Эти контекстные нарушения на уровне фраз, к удивлению самих исследователей, оказали влияние на динамику НР на довольно ранней стадии (рис. 2), но в то же самое время не привели к отклику, который можно было бы интерпретировать как N400. В одной из этих работ такая модуляция НР была обнаружена уже через ~115 мсек после того, как слова можно было различить; было найдено, что эта модуляция опосредована активностью в верхних височных и нижних лобных областях левого полушария (Shtyrov & Pulvermuller, 2007). В другой работе (Menning et al., 2005), контекстно-семантические аномалии нашли свое отражение в НР в диапазоне 150 - 200 мсек. Надо заметить, что эти показавшие контекстно-семантическую НР работы было проведены в разных лабораториях с использованием различного оборудования, протоколов стимуляции и языка стимулов (финский, немецкий). В этих работах впервые было показано, что семантические процессы высшего порядка — контекстной интеграции устной речи — могут происходить в рамках 200 мсек после предъявления необходимой информации, т. е. задолго до ранее известного феномена N400. Таким образом, вместе с рассмотренными выше данными фонетичеких, лексичексих и семантичексих работ, они предоставляют дополнительные доказательства в пользу раннего и параллельного доступа к разным типам лингвистической информации. Дополнительная поддержка этого взгляда на речевые процессы была также найдена и в опытах со зрительной стимуляцией (Sereno et al., 2003; Penolazzi et al., 2007), показавших эффекты контекстно-семантической интеграции на зрительные отклики мозга с латентностью менее 200 мсек.
Синтаксические процессыНаконец, давайте рассмотрим последний из основных типов лингвистических процессов, перечисленных нами ранее — обработку морфосинтаксической информации в мозге человека. Нами проведен ряд экспериментов по этой тематике, результаты которых также нашли свое подтверждение и в работах других лабораторий. Для обеспечения точного контроля над физическими свойствами раздражителей мы опять-таки предъявляли идентичные вербальные стимулы в различном контексте. В этих экспериментах критическое слово следовало за контекстным словом, которому оно либо соответствовало грамматически, либо находилось с ним в синтаксическом конфликте; это стандартный подход в нейрофизиологии синтаксиса (Osterhout, 1997). Использовались различные методики (МЭГ, ЭЭГ) и языки стимуляции (финский, немецкий, английский); это дает возможность делать общие выводы, что особо важно в экспериментах с НР, т. к. они обычно используют ограниченный набор стимулов. Первая из этих работ имела дело с нейрофизиологическими откликами на финские фразы, состоявшие из местоимения и глагола, имевшего либо верное в своем спряжении окончание (напр., «он идет», «ты идешь»), либо рассогласованного с местоимением в лице («он идешь», «ты идет»). Одни и те же стимулы использовались в синтаксически верных и аграмматических фразах; таким образом, влияние физических факторов было устранено. Результаты показали увеличение амплитуды НР в аграмматических контекстах по срав- нению с НР на те же слова, предъявляемые в грамматически верных фразах (рис. 2). Подобные результаты были получены в экспериментах с использованием английского и немецкого языков (Pulvermuller & Shtyrov, 2003; Pulvermuller & Assadollahi, 2007). Латентность, в которой регистрируются грамматические эффекты, различается в этих работах, но в целом они были найдены в пределах 200 мсек после пункта идентификации слова, а иногда и заметно раньше — около 100 мсек (Pulvermuller & Shtyrov, 2003; Shtyrov et al., 2003). Также разнятся и корковые структуры, в которых были найдены источники синтаксической НР. Магнитоэнцефалографические результаты указывают на распределение основных источников этой активности в верхне-височной области коры и на возможное участие нижнее-фронтальных областей, тогда как опыты с ЭЭГ указали на обратное, сильную активацию левых нижне-фронтальных областей вкупе со слабым участием височной коры. Эти различия можно отметить и в предыдущих работах по картированию синтаксиса в мозге, в которых эта функция локализуется иногда в лобных, а иногда в височных долях (напр., Kaan & Swaab, 2002; Bornkessel et al., 2005; Kircher et al., 2005). Таким образом, возможно, что в грамматико-синтаксические процессы вносят свой вклад различные области персильвиева неокортекса, и что их активность находит различное отражение в ЭЭГ и МЭГ, которые, как известно, отличаются по своей чувствительности к различному положению и направлению источников тока в мозге. Наши данные были также подтверждены работами других исследователей, которые при использовании схожих методов обнаружили специфическую НР с пиком в диапазоне 150 - 200 мсек при предъявлении фраз содержащих синтаксический конфликт (напр., Menning et al., 2005; Hasting et al., 2007). Ранняя синтаксическая НР напоминает компонент ELAN (Neville et al., 1991; Friederici et al., 1993), ранее предложенный в качестве индекса построения мозгом синтаксической структуры фразы (Friederici, 2002). Эта интерпретация поддерживается и данными о синтаксической НР. Не менее важно то, что они также указывают на то, что такие ранние синтаксические процессы не требуют внимания к речевой информации. В этом плане ранняя синтаксическая обработка представляется достаточно автоматическим процессом. Этот автоматизм проявляется даже в описанном выше случае нарушения согласования между подлежащим и спряжением сказуемого, которое является более ресурсоемким с точки зрения вычисления зависимостей и поэтому неспособным, как считалось прежде, к вызову ELAN. Более поздний сдвиг положительной полярности (P600), как оказалось, полностью отсутствует в пассивной парадигме рассогласования, и в свою очередь, видимо, отражает вторичные контролируемые (с участием внимания) попытки интерпретации последовательности после того, как первичный автоматический парсинг не удался (Osterhout & Holcomb, 1992; Friederici, 2002).
Отсутствие внимания и автоматизмПредставляется вполне возможным, что обычное требование к испытуемым сосредоточиться на просмотре фильма или чтении книги во время слуховой стимуляции не гарантирует полного отсутствия внимания к раздражителям. Обычно делаемый из таких экспериментов вывод об автоматизме изучаемых процессов подразумевает, что испытуемые строго следуют данной инструкции, в чем трудно полностью убедиться. Таким образом, предпочтительным представляется более строгий контроль над уровнем внимания испытуемых к стимуляции. В связи с этим нами был проведен эксперимент по более подробному исследованию роли внимания в языковой функции. В нем мы сравнили классическую парадигму НР с ее традиционно умеренным уровнем отвлечения испытуемых от стимуляции при помощи просмотра видеофильма и более требовательную задачу для испытуемых, в которой от них требовалось непрерывное выполнение задания по классификации альтернативных слуховых стимулов. Речевые стимулы проигрывались только в правое ухо, в то время как неречевые акустические раздражители, требовавшие определения, подавались в левое ухо. В одном из экспериментальных блоков испытуемым было необходимо внимательно слушать неречевую симуляцию в левом слуховом канале и нажимать кнопку всякий раз, когда там появлялся девиантный сигнал; в то же самое время, языковая стимуляция поступала в правое ухо. В другом блоке, испытуемым была дана инструкция смотреть видеофильм, как в большинстве предыдущих работ, в то время как оба вида стимулов предъявлялись каждый в своем канале, но без задания уделять им какое-либо внимание. При использовании синтаксических языковых стимулов, мы снова получили грамматический эффект, т. е. более сильный отклик НР в ответ на грамматически неверные сочетания по сравнению с правильными фразами. В диапазоне времени до 150 мсек не было обнаружено достоверных различий между задачами с различным уровнем отвлечения внимания (рис. 2). Лишь позднее, после 150 мсек, мы обнаружили статистически достоверное взаимодействие между экспериментальными факторами, указавшее на то, что эти более поздние стадии обработки грамматической информации мозгом, находящие свое проявление в НР, подвержены влиянию внимания и типа экспериментального задания. Наша интерпретация данных результатов заключается в выводе о независимости ранней части НР от внимания и автоматическом характере раннего синтаксического анализа (Pulvermuller et al., 2008).
Заключение: выводы и нерешенные вопросыВ настоящем обзоре мы рассмотрели предпосылки к нейрофизиологическому исследованию самых ранних стадий восприятия языка мозгом и то, как для их обнаружения и исследования можно использовать негативность рассогласования. Важными предпосылками для такого использования НР нам представляются ранний характер НР, автоматизм ее генерации, возможность записи откликов на индивидуальные языковые стимулы в парадигме рассогласования и уникальная характеристика НР как отклика на акустический контраст, что позволяет осуществлять дополнительный контроль над физическими свойствами экспериментальных раздражителей. Мы увидели, как НР использовалась в последние годы для исследования нервных процессов, связанных с обработкой фонологической, лексической, семантической (включая семантику на уровне отдельных слов и на более высоком уровне контекстной интеграции фраз) и синтаксической информации мозгом. НР не только обнаружила чувствительность к этим типам лингвистической информации; более того, она оказалась полезной для выделения мозговых коррелятов лингвистической обработки из общей массы процессов, связанных также со вниманием и стратегией анализа раздражителей. Не менее важным представляется то, что работы с использованием НР дали возможность строго контролировать свойства раздражителей, минимизировав таким образом возможность того, что обнаруженные эффекты вызваны акустически или психолингвистически несбалансированными наборами стимулов. Важно также и то, что эти работы пролили новый свет на временную динамику доступа к лингвистической информации. В различных экспериментах различные типы стимулов включая фонетические/фонологические, лексические, семантические и синтаксические контрасты, модулировали НР в диапазоне латентности 100?200 мсек после того, как соответствующая акустическая информация становилась доступна на входе в слуховую систему. Эти данные недвусмысленно говорят в пользу параллельного или приблизительно одновременного доступа к различным типам лингвистической информации, быстро происходящего вскоре после того, как услышанные слова можно идентифицировать. Такой доступ был ранее предложен некоторыми психолингвистическими исследованиями, но по сию пору являлся предметом острых теоретических дискуссий. В структурном плане, полученные с помощью НР результаты четко указывают на существование распределенных нейронных сетей, лежащих в основании обработки поступающей речевой информации. Например, был показан комплексный характер активации верхне-височных и нижних лобных областей коры в процессе восприятия слова. В зависимости от конкретных семантических свойств предъявляемых слов, они приводят к активации целого набора областей в височной, нижне-фронтальной, латерально- и дорзофронтальной коре, иногда распространяющейся и в правое полушарие (Pulvermuller al., 2004). Предоставляемые НР возможности для исследования таких сложных пространственно-временных паттернов, связанных с восприятием и пониманием языка очевидно, далеко не исчерпаны на данном этапе. Полученные с помощью НР результаты ни в коем случае не отрицают более ранние работы по определению нейрофизиологических параметров лингвистических процессов. Скорее, они придают новое измерение нейрофизиологии языка, указывая на то, что более поздние процессы, находящие свое отражение, напр., в N400, P600 или синтаксических негативностях (LAN), могут являться дополнением и продолжением ранних автоматических этапов и таким образом, могут быть связаны со вторичными умственными процессами, запускаемыми ранним доступом к лингвистическим следам памяти и зависимыми от внимания и индивидуальных стратегий. Исследования языковых процессов с помощью НР имеют, тем не менее, и ряд нерешенных вопросов. Одним из наиболее часто подвергающихся критике моментов такого подхода является контроль над вниманием в экспериментах с НР. Традиционные пассивные парадигмы НР, в которых основной задачей испытуемого является просмотр фильма, чтение книги или компьютерная игра, в действительности не могут исключить возможность того, что испытуемый может время от времени «подслушивать» экспериментальные стимулы (Carlyon, 2004). Поэтому одним из направлений дальнейшего развития этих исследований является дальнейшее усовершенствование экспериментального протокола, который должен гарантировать более надежное отвлечение внимания от стимуляции. Последнее может включать в себя отвлекающие задачи, предъявляемые в той же самой модальности, для разделения независимых от внимания и потому возможно автоматических лингвистических процессов от тех, которые требуют активного участия индивидуума. Как уже указывалось, мы с успехом использовали такой подход для подтверждения независимости синтаксической НР от внимания на ее самых ранних стадиях. Представляется необходимым перенести этот подход на исследования различных типов лингвистической информации. Другим критическим моментом подхода НР к языку является небольшое количество стимулов, которое можно использовать в контексте одного эксперимента (достоинства этого подхода мы рассмотрели выше). Малое число используемых раздражителей и связанная с этим необходимость их многократного повторения для записи качественного ВП, безусловно, приводят к трудностям в обобщении любых полученных результатов на весь естественный языковой ландшафт. В данном случае, несомненно полезными оказались повторные опыты с использованием различных слов, языков и экспериментальных установок. Возможным будущим направлением представляется увеличение разнообразия стимуляции, делая ее более репрезентативной по отношению к естественной речи. Такая стратегия уже успешно применялась к отдельным фонемам (Shestakova et al., 2004). Однако, перенос этого подхода на более сложные стимулы, хотя бы на уровень слов, требует строгого контроля над их физическими и, не менее важно, психолингвистическими параметрами, а также над вариативностью этих мер в группах экспериментальных стимулов. Несомненно, такая задача окажется нетривиальной для исследователей, желающих взяться за ее решение. И, наконец, имеет место очевидный конфликт между, с одной стороны, традиционной интерпретацией НР, которая связывает ее с механизмами определения изменений в слуховом сенсорном канале и процессами кратковременной памяти, и, с другой стороны, природой откликов НР на речевые стимулы. Последние требуют для своего появления наличия следов долговременной памяти, а также проявляют сложные пространственно-временные паттерны, зачастую распространяющиеся на области за пределами слуховой коры. Это привело к предположению, что лингвистическая активация в парадигме рассогласования является отдельным процессом, который происходит в дополнение к классической, связанной с детекцией изменений негативности рассогласования (Pulvermuller & Shtyrov, 2006). Теоретически, можно предположить, что такие процессы не обязательно требуют затратной в плане длины эксперимента парадигмы рассогласования и, возможно, могут быть зарегистрированы и в других экспериментальных условиях. Таким образом, дальнейшее изучение экспериментальных условий, необходимых для надежной регистрации активации долговременных следов памяти в мозге в отсутствие внимания и при строгом контроле над параметрами стимулов, может не только пролить свет на лежащие в их основе процессы, но и привести к появлению улучшенных экспериментальных парадигм для исследования мозговых основ языковой функции человека. Список литературы Alho, K. (1992) Selective attention in auditory processing as refected by event-related brain potentials. Psychophysiology, 29, 247?263. Alho, K. (1995) Cerebral generators of mismatch negativity (MMN) and its magnetic counterpart (MMNm) elicited by sound changes. Ear Hear, 16, 38?51. Bentin, S., Mouchetant-Rostaing, Y., Giard, M. H., Echallier, J.F. & Pernier, J. (1999) ERP manifestations of processing printed words at diferent psycholinguistic levels: time course and scalp distribution. J Cogn Neurosci, 11, 235?260.
Комментарий
Слова представлены в нервной системе в виде специфических следов памяти, существующих как распределенные сети нейронов. Распределенные сети нейронов (РСН) формируются в результате усиления двусторонних связей между различными (в т.ч. удаленными друг от друга) областями мозга по мере того, как индивидуум получает опыт взаимодействия с движениями, объектами и концепциями одновременно с описывающими их словами (Hebb, 1949; Pulvermuller, 1999). Такая связь слова с его конкретным значением является неотъемлемой частью его семантики (Frege, 1980). Таким образом, выводы обзора Штырова Ю. Ю. и Пулфермюллер Ф., соответствуют предположениям, что лингвистическая деятельность осуществляется мозгом в значительной степени автоматизированным образом (Fodor, 1983; Garrett, 1984; Garrod & Pickering, 2004; Pickering & Garrod, 2004). По сути, все эти исследования указывают на наличие информационной системы распределенных нейронных сетей, которая реализует обработку речевой информации автоматическим образом вне фокуса внимания. Системный подход – предусматривает рассмотрение объектов как систем; ориентирует исследователя на раскрытие целостности объекта, на выявление многообразных типов связей в нем и сведение их в единую теоретическую картину. Науку интересует выделение существенного для решения тех или иных задач в «чистом» виде, т. е. в отвлечении ото всего несущественного. И этого нельзя добиться без идеализации и моделирования. Идеализация играет огромную роль в науке. Например, теории описывают не предметы «сами по себе», а их идеализации, которые применяются для выявления существенного в этих предметах для решения конкретных задач, поставленных наукой. Поэтому можно сформулировать принцип идеализации. Суть этого принципа состоит в том, что для выделения существенного необходима идеализация того объекта, который подлежит изучению. При этом сущность есть основные признаки объекта, обусловливающие все остальные его признаки. Этот принцип говорит об относительности идеализаций применительно к решаемым с их помощью задачам.
Рассмотрим идеализации элементов информационной системы распределенных сетей нейронов (РСН) мозга учитывая, что вербальные определения всегда идеализируют объект.
Элементы информационной системы РСН
Определение мысли на основе информационной системы РСН
Мысль - 1) элементарная мысль – выявленная в сознании ассоциативная связь между 2 - 3 элементами информационной системы РСН: образом субъекта, образом процесса, образом объекта, образом свойств, которая имеет место в реальной ситуации или может быть исходя из сознания конкретной и воображаемой ситуации. При замене этих элементов информационной системы РСН на описывающих их слова формируется исходная фраза, соответствующая конкретной и воображаемой ситуации. 2) Элементарная мысль дополнительно с логической функцией информационной системы РСН – понятие, в которой обобщены и выделены объекты, процессы, признаки некоторого класса по определенным и в совокупности специфическим для них признакам.
Практическая реализация элементарной мысли
Исходная фраза первоначально формируется и соответствует характеристикам второго этапа развития речи ребенка (1,5 – 2,5 года). Может соответствовать предложению. Исходная фраза – единица речи, выражающая законченную мысль, которая является основной элементарной мыслью в данном предложении и в соответствии с авторским замыслом преобразуется (дополняется) в авторскую последовательность самостоятельных словосочетаний (фраз) используемых в сложном предложении (подробнее). Теперь мы можем конкретно изучать и исследовать процессы возникновения и развития человеческой речи, а именно процессы формирования информационной системы распределенных нейронов мозга. Т.е. могут быть уточнены и скорректированы известные теоретические проблемы возникновения и развития речи (подробнее).
Эта статья – одно из необходимых звеньев, которые позволили, используя системный мультидисциплинарный подход, ответить на ключевые вопросы:
1. Психология (подробнее)
До сих пор перед психологами стояли ключевые вопросы, на которые они не могли ответить:
Эти вопросы оставались предметом жарких споров, когда философы по-прежнему были более сведущи в этих вопросах, чем психологи (Глейтман, 2001).
2. Молекулярная генетика (подробнее)Эволюция человека сопровождалась изменением активности генов-регуляторов. Почти все гены, чья активность в клетках мозга сильно различается у человека и шимпанзе, у человека работают активнее. Что бы это значило? Ранее было неясно...
3. Психолингвистика (подробнее)Для конкретного ключевого события в культурной эволюции человечества стало возможным определить конкретный этап развития речи и его содержание.
В соответствии с особенностями формирования информационной системы распределенных сетей нейронов (РСН) мозга и функций субъективного мира человека (СМЧ) пятого уровня (подробнее) мы можем утверждать, что речь первобытных художников расписавших пещеру
Появление лука, как ручного оружия для метания стрел, позволяет выявить более поздние даты, связанные с обработкой лингвистической информации, соответствующей последующей стадии развития речи ребенка с 4,5 лет до 6 – 7 лет (подробнее). Даты ключевых событий в культурной эволюции человека, в его «духовном развитии» связаны с механизмом регуляции экспрессии генов (подробнее).
Дата публикации: 16.06. 2012г. 02:30 Юрий Павлов
|
|

 (на сегодняшний день это - самая ранняя на Земле живопись - около 32 тыс. лет), соответствует стадии развития речи
ребенка, которая начинается с 3,5 лет и продолжается до 4,5 лет.
(на сегодняшний день это - самая ранняя на Земле живопись - около 32 тыс. лет), соответствует стадии развития речи
ребенка, которая начинается с 3,5 лет и продолжается до 4,5 лет.